
I take a lot of photographs. Somewhere between playing my C. Bechstein B-212 grand piano, and golf, high-fidelity photography is one of the hobbies I keep returning to, and over the years, I have developed a deep respect for two very different machines.
On one side, there is the Hasselblad. A medium-format Hasselblad produces images of almost absurd fidelity — enormous sensors, exquisite glass, resolution, and tonal depth that an entire genre of professional photographers will never give up. When you want to capture a single, still subject in all its glory — a face, a landscape, a product on a table — nothing touches it. It is the gold standard for a perfect frame.

On the other side, there is the phone in my pocket. By any classical measure, an iPhone sensor is tiny, and its lens is humble. And yet it has quietly become the most consequential camera on Earth. Not because it out-resolves a Hasselblad — it does not — but because it does something a Hasselblad was never designed to do. It fuses dozens of frames in real time, it shoots video, it captures the laugh, the jump, the wave, the moment. It trades a little resolution for an honest record of what moves.

I keep coming back to this contrast, because it is the cleanest way I know to explain what my field has gotten wrong about proteins — and what a paper we just published is trying to fix.
For most of the modern history of structural biology, our ambition was Hasselblad ambition. We wanted the perfect still.
We crystallized a protein, hit it with X-rays, and reconstructed a single, gorgeous, atom-by-atom structure. We learned a tidy story along with it: sequence folds into structure, and structure dictates function.

Find the one true shape, find the pocket, design the drug. The whole edifice of classical, structure-based drug discovery is, in a sense, a Hasselblad pointed at biology — an instrument optimized to capture one immaculate, motionless frame at the highest possible resolution.

And for proteins that actually hold still, it is glorious. A well-folded enzyme sitting in its crystal is the perfect Hasselblad subject: posed, static, cooperative. Click. You get a masterpiece.
And we did not just take these portraits one at a time. We built an archive.
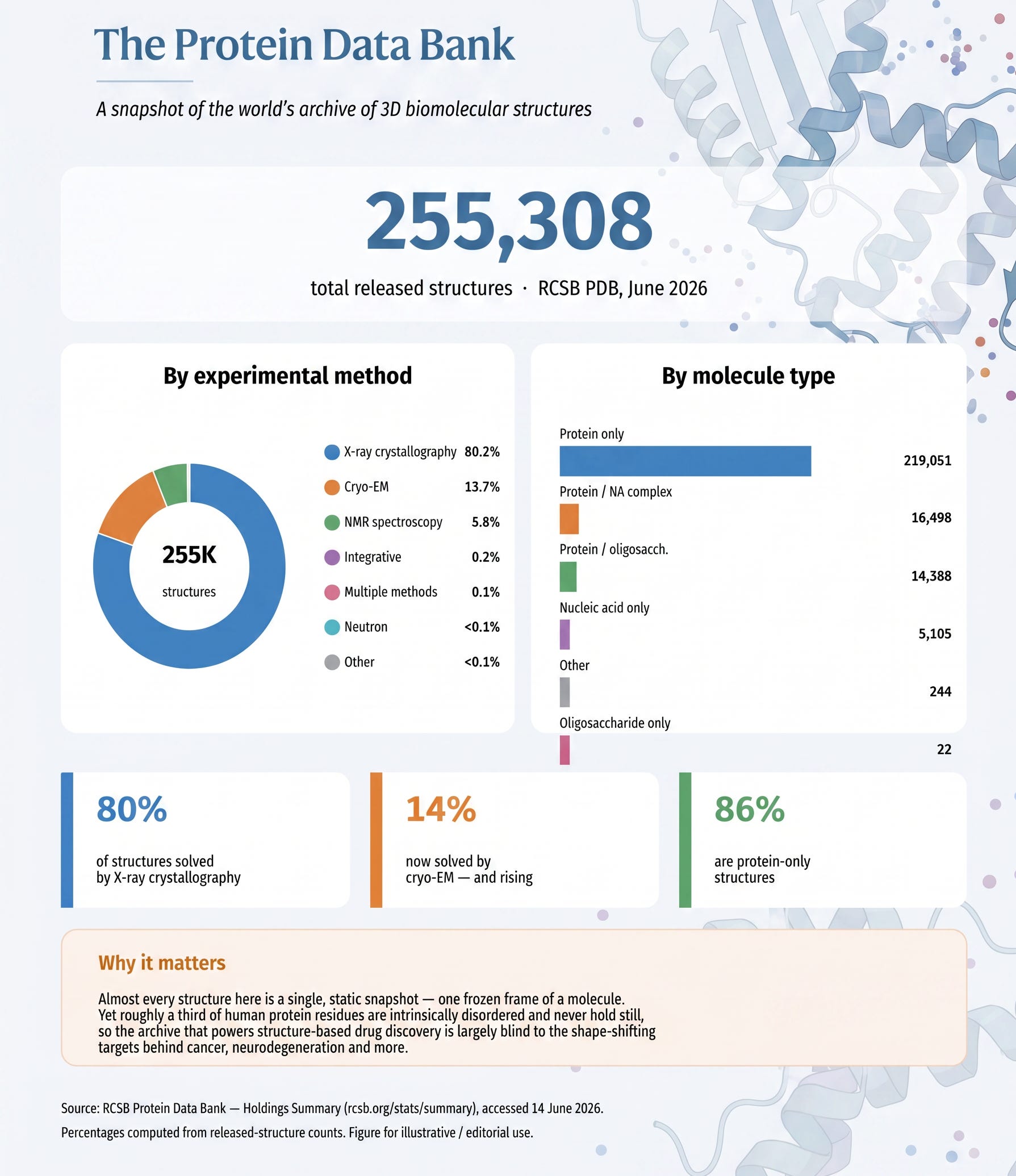
The Protein Data Bank is the great gallery of structural biology — a single, public repository where, for more than fifty years, every beautiful still we captured has been hung on the wall for everyone to study. Today it holds over 255,000 structures, the overwhelming majority of them solved by exactly the kind of Hasselblad instruments I have been describing: around four out of five by X-ray crystallography, with most of the rest from cryo-EM and NMR (RCSB Protein Data Bank). It is one of the most extraordinary collective achievements in all of science — a quarter of a million frozen frames of life’s machinery, each one painstakingly earned.
And here is where the story takes its most thrilling turn. Because once you have a gallery that large, you can do something new with it: you can teach a machine to paint.
That is, in essence, what AlphaFold did. DeepMind’s AlphaFold2, and now AlphaFold3, learned to predict a protein’s three-dimensional structure straight from its sequence — and they learned it by studying the Protein Data Bank. AlphaFold3 is explicitly built to predict “nearly all molecular types present in the Protein Data Bank,” trained on the very structures deposited there over decades (Abramson et al., Nature, 2024).

It was a staggering achievement, and it kicked off an open-source renaissance: OpenFold faithfully retrained the AlphaFold2 architecture from scratch on roughly 132,000 experimental PDB structures (Ahdritz et al., Nature Methods, 2024), while Chai-1 and Boltz reproduced AlphaFold3-level accuracy and released their weights to everyone — each of them, again, trained on the full PDB (Boltz-1, MIT Jameel Clinic). In just a few years, we went from one frozen frame at a time to models that conjure a plausible frame for almost any sequence we hand them, in seconds.
But notice what every one of these remarkable machines inherited from its teacher. They learned from a gallery of stills. They are, all of them, the most sophisticated Hasselblads ever built — trained on perfect portraits to produce another perfect portrait. Ask AlphaFold for a structure, and it will hand you one gorgeous, confident, motionless frame, because that is the only kind of picture its entire education ever contained.

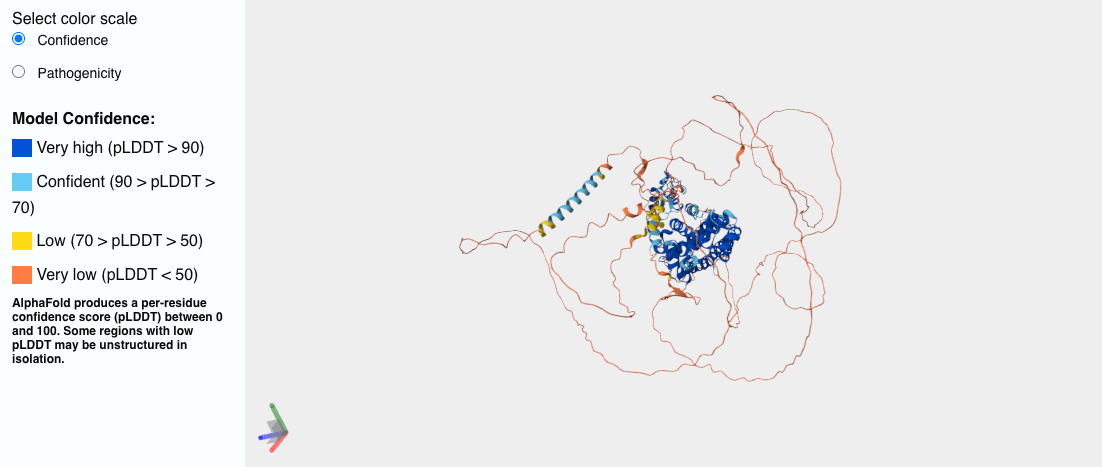
For a protein that holds still, this is pure magic. For one that doesn’t, for example, human Androgen Receptor (AR), the key driver of prostate cancer, the machine simply paints you the most convincing still it can of a subject that never sits still in the first place — and hands it over with a confidence that hides the very thing you needed to know. Just see the image below, the ‘spaghetti’ parts of the protein are all red and, from a structural biochemistry and drug discovery perspective, unusable.
Asking a Hasselblad to photograph an intrinsically disordered protein is like asking it to photograph a hummingbird in flight. You will get one technically flawless frame of a blur — and you will learn almost nothing about how the bird actually flies.

This is the part that took me years to fully internalize, so let me say it plainly.
For a disordered protein, a single high-resolution structure is not just insufficient — it can be actively misleading. It implies a permanence that does not exist. It is a photograph of a thing pretending to be something it is not. You walk away with a stunning image and a false belief: that the molecule looks like that.
It does not look like that. It looks like that, and that, and that — a shifting ensemble of interconverting shapes, most of them fleeting, visited and abandoned over and over. The function lives in the motion, not in any one frame, just like in the attached example of the ACTR protein.
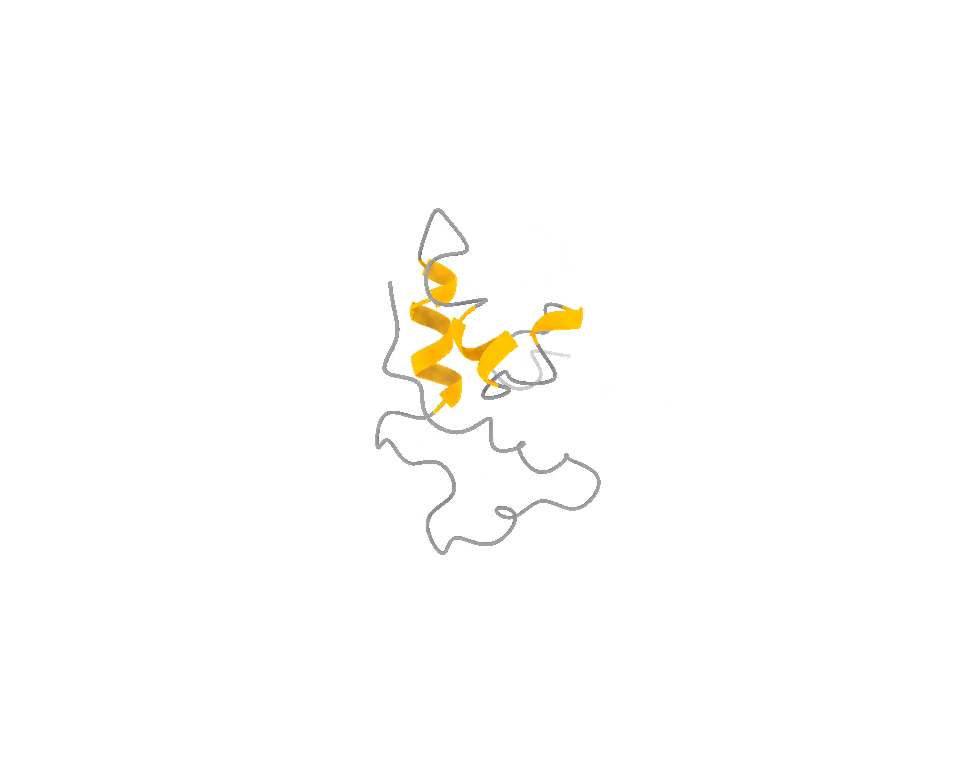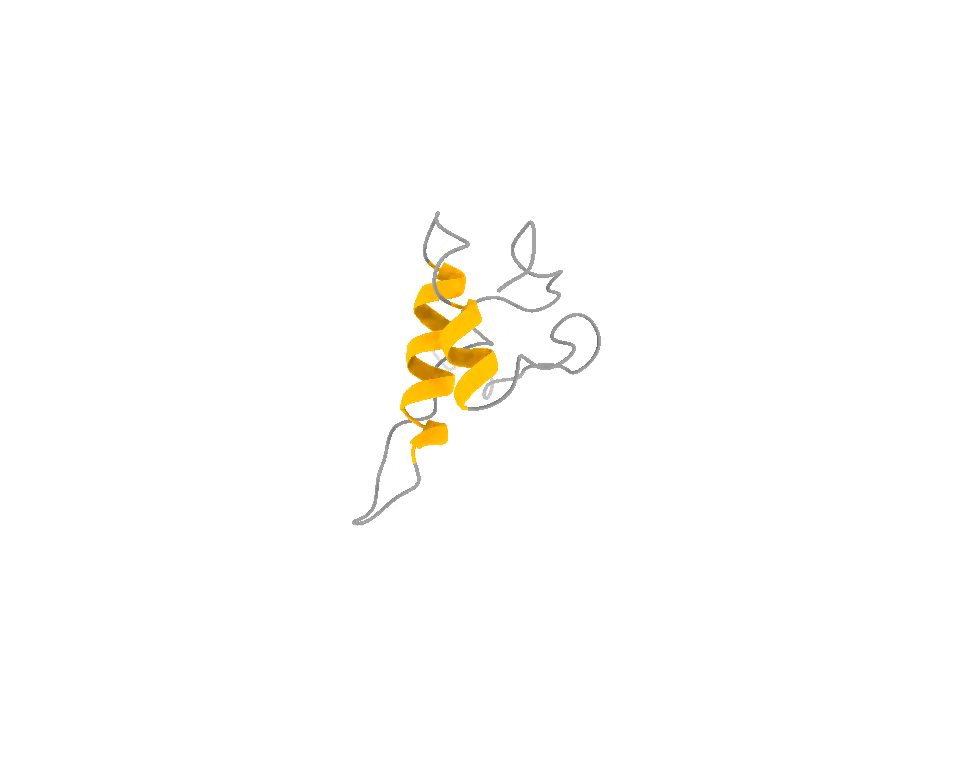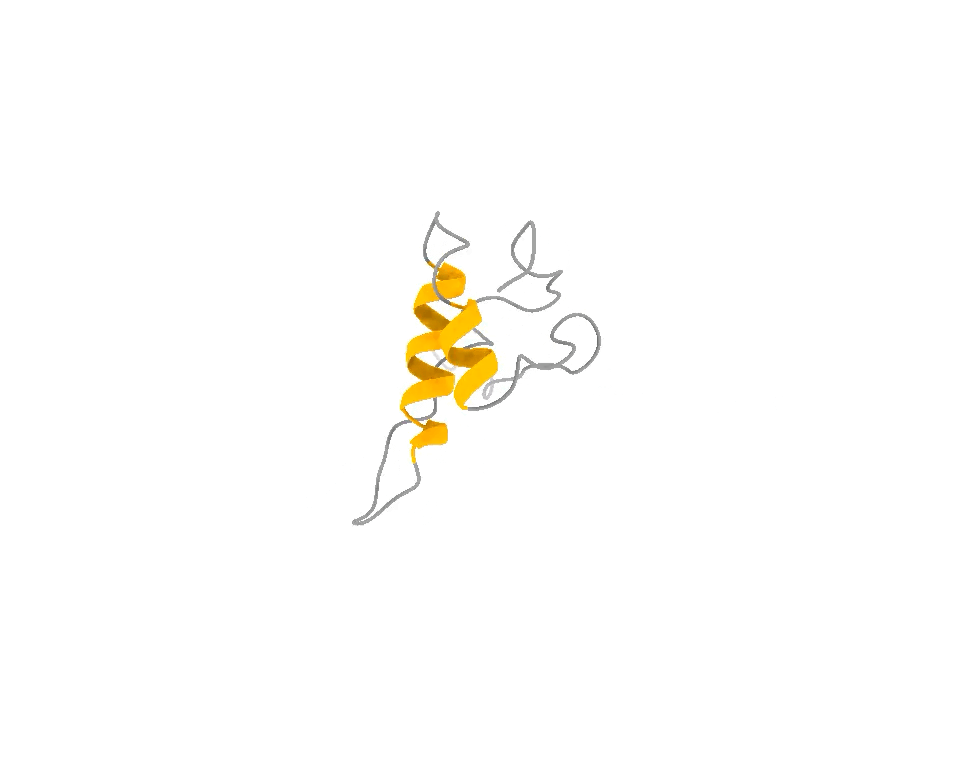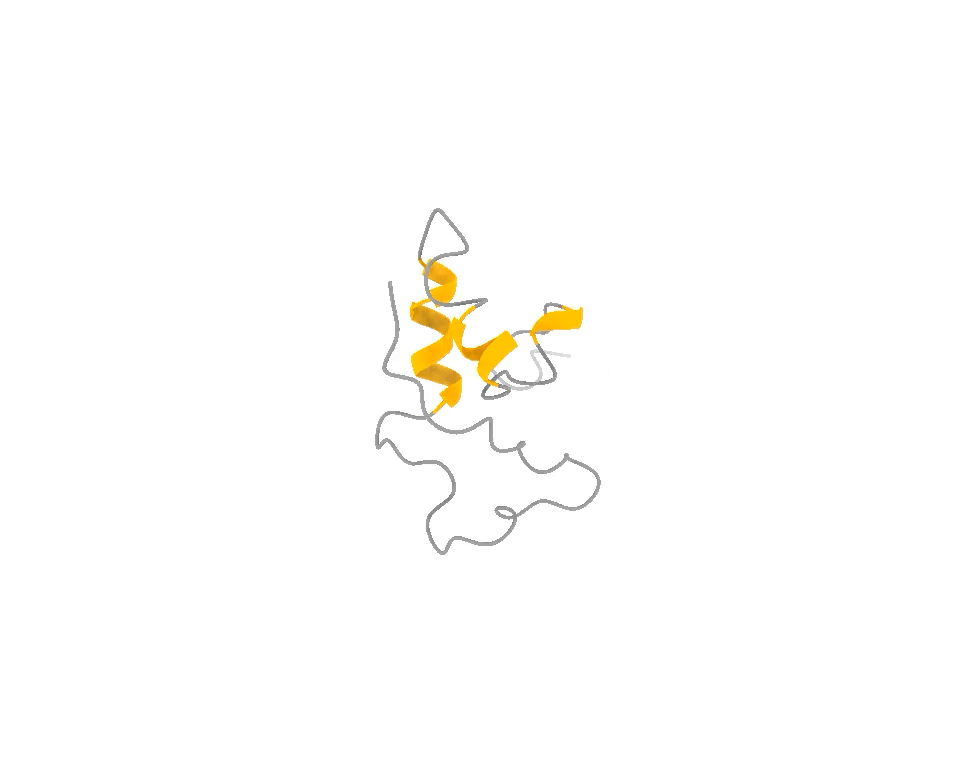
So you face a genuine trade-off, and it is exactly the photographer’s trade-off. You can have one frame at the highest possible resolution and lose all the motion. Or you can accept slightly lower per-frame resolution and, in exchange, capture the thing that actually matters: the dynamics, the rare moments, the fleeting shapes the molecule briefly assumes when it is doing its job. For a hummingbird — and for a disordered protein — the second option is not a compromise. It is the only honest picture.
A Hasselblad gives you a singular state at high resolution. An iPhone gives you dynamics at sensible fidelity. For proteins that refuse to hold still, the iPhone wins — not despite the trade-off, but because of it.
Here is the detail I love most, because it maps so cleanly onto our work.
The iPhone does not beat physics. Its sensor really is small. What makes it extraordinary is computation — it captures many frames, understands how they relate in time, and fuses them into something richer than any single exposure could ever be. The intelligence is not in the glass. It is in how the frames are sampled and combined.

That is almost exactly what we set out to do with disordered proteins, except our “camera” is a physics simulation.
In our new paper in Nature Communications, my colleagues applied an enhanced-sampling method called On-the-fly Probability Enhanced Sampling — OPES — in a multithermal ensemble.

The technical achievement is that a single simulation replica diffuses across a wide temperature range and is reweighted back to physiological conditions, instead of running the brittle, parameter-heavy replica-exchange machinery these simulations normally demand (Streit et al., 2026). Think of it as one clever, computationally fused exposure rather than a cumbersome rig of cameras all firing at once.

And the OPES “footage” holds up against reality. Compared to the older REST2 approach and to plain simulations, it produces consistent ensemble averages, converges faster, and — crucially — explores broader conformational space, catching low-population states the other methods miss (Streit et al., 2026). Lower drama, more honest motion.
We pointed this iPhone-style camera at ACTR, a 71-residue disordered transcriptional coactivator that only folds when it meets its partner. In its free state, ACTR looks like exactly what the textbooks promise: a floppy, structureless chain. Photograph it with a Hasselblad and you would conclude there is nothing there to see.
But the motion told a different story. Our simulations revealed a rare, low-population set of transiently structured states — populated only a few percent of the time — in which multiple α-helices fold cooperatively and form genuine tertiary contacts, a fleeting, binding-competent shape the protein visits and then lets go (Streit et al., 2026). This is the laugh in the candid photo. The wave. The moment a posed portrait would never have caught.
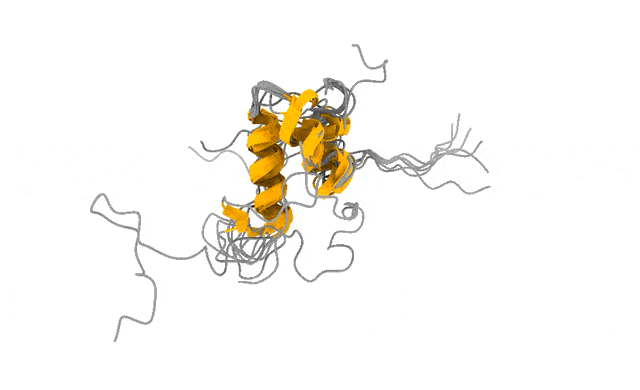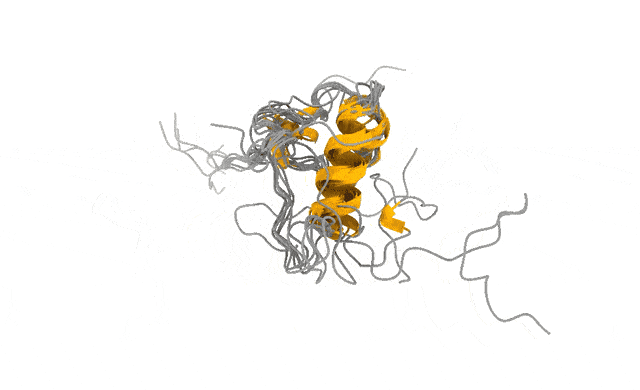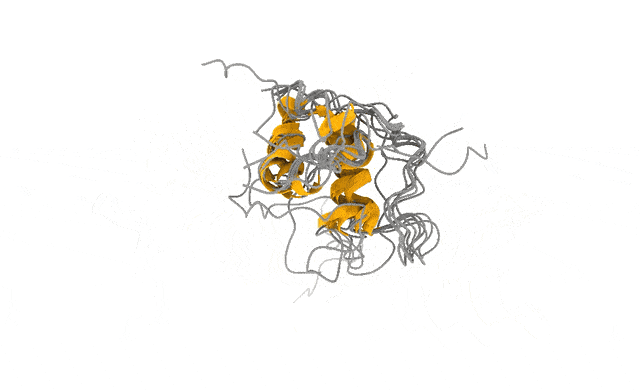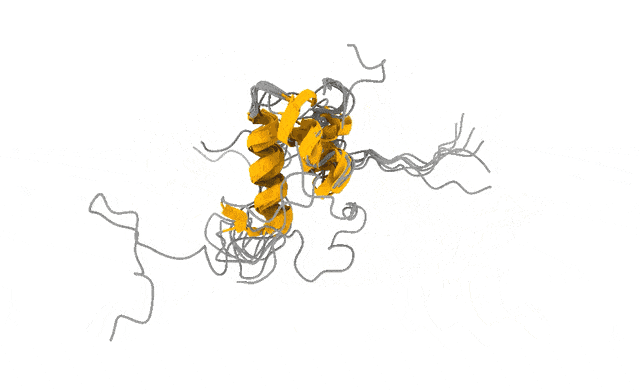
And these are not numerical ghosts. The states are reversibly sampled, separated from the disordered ground state by only a modest free-energy barrier, and — the part that matters most to me — consistent with extensive NMR and SAXS data we used to validate the ensemble (Streit et al., 2026). The physics and the experiments agree on what the candid shot shows.
Why should anyone outside a biophysics lab care?
Because those fleeting, partially folded conformations may harbor exactly the kind of binding pockets that make an “undruggable” disordered protein suddenly addressable.
You cannot design a drug against a pocket you have never seen. And you will never see this one with a Hasselblad.
None of this means the Hasselblad is obsolete. For a folded enzyme with a deep, stable pocket, the perfect high-resolution still is still the right tool, and I would reach for it every time. The mistake was never owning a Hasselblad. The mistake was believing it was the only camera worth owning — and pointing it at subjects it was never built to capture.
The lesson photography taught me, and the lesson this paper makes concrete, is the same one:
Resolution is not the same as understanding.
A slightly lower-resolution picture that captures motion can tell you infinitely more than a flawless picture of a single frozen instant — when the truth of your subject is the motion.

For a third of the proteins in your body, the truth is the motion. Just like the human body, the proteins we care about are always in motion. So we stopped trying to take the perfect photograph and started shooting the honest, computationally fused, slightly lower-resolution footage instead. It is the least glamorous-sounding upgrade in the world. Trade a little resolution, catch the moment.
Big thanks to Kareem Reeda for reading, reviewing, and providing critical suggestions.
I want to cordially thank our Scientific Advisory Board member, Prof. Kresten Lindorff-Larsen of the University of Copenhagen, for his continued scientific support and for spearheading our efforts to advance understanding of disordered proteins.
I want to express my sincere gratitude to Dr. Julian Streit, who conducted this project jointly with our Peptone team: Dr. Michele Invernizi and Dr. Sandro Bottaro.