о холоднокровности пчелы
Помню, в школе на уроках биологии рассказывали про теплокровных и холоднокровных животных; насекомых, а, значит, и пчелу относили к холоднокровным. Поэтому когда первый раз прочитал, что пчела - пойкилотермное животное, слегка насторожился. Оказалось - зря, это тоже, что холоднокровное, но “по-научному”, ещё встречается “эктотермное”. А теплокровные - гомойтермные или эндотермные; они способны сохранять постоянную температуру тела, независимо от температуры окружающей среды - это птицы и млекопитающие, остальные - холоднокровные. Итак, согласимся: пчела - пойкилотермное животное.
Однако, “всё не так однозначно”. Например голый землекоп - холоднокровное млекопитающее. А в мае 2015 года нашли “полностью теплокровную рыбу”. Оказалось, правда, на мой взгляд, не полностью - она способна держать температуру всего на 5°С выше окружающей. Выделяют отдельную группу гетеротермных животных, куда относят как некоторых холоднокровных, так и некоторых теплокровных - они могут менять интенсивность собственной теплопродукции. К ним относят насекомоядных грызунов, летучих мышей, медведей. Гетеротермия свойственна также ряду мелких птиц с быстрым обменом веществ, с её помощью переживающих ночное время, а также периоды бескормицы. Так, стрижи в дождливую погоду способны голодать до 4-х дней, при этом температура их тела падает до 20°C. Цепенеют на ночь и колибри, температура их тела снижается при этом до 20-17°C. Среди холоднокровных гетеротермия встречается у некоторых летающих насекомых, рыб, питонов. В определённом состоянии температура их тела может значительно превышать температуру среды (например, у насекомых во время полёта).
Ещё гомойтермию делят на истинную и инерциальную. При инерциальной - температура тела, нагретого за день на солнце, не успевает упасть за ночь за счёт большой массы и размеров. Этим характеризуются крупные крокодилы, черепахи, вараны, питоны, удавы.
Медоносная пчела в отличие от многих холоднокровных не впадает в спячку в зимнее время, способна поднять свою температуру не сходя с места, а в полёте она у неё может быть на 15°С выше окружающей, имеет волосяной покров, что характерно именно для теплокровных. Что-то не очень похоже на холоднокровность…
Знаменитый Bernd Heinrich в 1993 году даже издал книгу под названием “Теплокровные насекомые. Стратегии и механизмы терморегуляции” The Hot-Blooded Insects. Strategies and Mechanisms of Thermoregulation.
Профессор Т. Цесельский более века назад (книга 1913 года второе издание перевода с польского) тоже усомнился в верности отнесения пчелы к холоднокровным и решил измерить температуру внутри пчелы своими руками:

Мы этого делать (размозжать в ступке - “O tempora, o mores!”) не будем и ограничимся уже имеющимися опубликованными данными. И, да, признаем, что сомнения профессора в холоднокровности пчелы обоснованы.
В общем, дело в терминологии. По мере накопления новых фактов возникает определённая кластеризация, и для её описания вводят термины. Однако, как видим, имеет место нечёткая логика. Поэтому не будем обращать внимание на категории и имеющиеся классификации - “хоть горшком назови, только в печку не ставь!” - а просто рассмотрим, как меняется температура и теплопродукция пчелы в зависимости от внешних условий.
В процессе метаболизма в организме пчелы из пищи (мёд и перга) извлекается энергия. Она расходуется на обеспечение жизнедеятельности организма, выработку необходимых ферментов и пр. и механическую работу мышц - мышц сердца для прокачки гемолимфы, мышц ног для перемещения по твёрдой поверхности, мышц крыльев для совершения полёта, а также мышц брюшка, мышц головы и органов головы и др. Энергия химических связей преобразуется в механическую с определённым коэффициентом (КПД), оставшаяся часть переходит в тепловую форму, повышая температуру тела пчелы и рассеиваясь в пространство. Скорость метаболизма и соответствующая ей мощность при этом разная для разных процессов - в покое, в полёте, в улье, в клубе. Разным будет и тепловыделение, называемое обычно термогенезом.
В покое. Итерация 0.
В начале своего развития от яйца к личинке и куколке пчела находится в инкубационных условиях при стабильной температуре, поддерживаемой семьёй с высокой точностью в районе 35-36°С. Видимо, эта температура оптимальна и для взрослой пчелы. Пчела может нормально существовать (жить) в интервале температур окружающего воздуха от 14 до 39°С. За пределами этого интервала, она тоже может жить, но недолго. В этом случае она стремится покинуть неблагоприятную среду или, объединяясь с другими пчёлами, предпринимает меры к нагреванию или охлаждению среды и друг друга до приемлемой температуры - создаёт локальный микроклимат. Для создания математической модели нужны данные. Их оказалось не так много. В покое температура тела пчелы выше температуры окружающего воздуха на 9°С при 10°С и на 1°С при 34°[1]. Т.е. при 10°С температура пчелы 19°С, а при 34°С - 35°С. Вот и все данные. А что между этими точками или за пределами этого интервала? Ничего не остается, как предположить, что между ними - прямая линия, по двум точкам больше ничего не построишь.
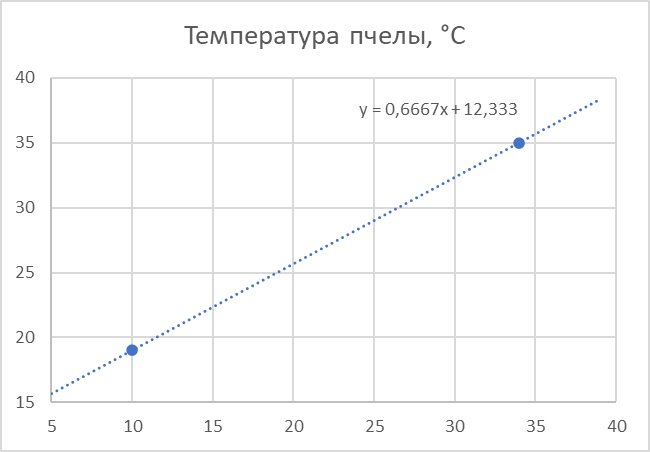
Еще данные (опять две точки): при 30°С пчелы активно двигаются, потребление кислорода при этом составляет 1,13-1,15 мл/пчелу в час, при 20 °С большинство из них неподвижны, но потребление кислорода составляет 2,25-2,45 мл/пчелу в час [2]. Если перевести объем кислорода в массу, то получим, соответственно, 1,65 мг/пчелу в час при 30°С и 3,3 мг/пчелу в час при 20°С. Известно, что кислород расходуется в количестве 0.86 кг на каждый килограмм мёда, или 1.16 мг меда сжигается на 1 мг кислорода. Отсюда можно посчитать, сколько меда сжигает пчела в час при разных температурах, а зная калорийность меда, определить какая при этом выделяется энергия. Получится на 1 мг кислорода в час пчела генерирует 4.2 мВт. Соответственно, при 30°С мощность термогенеза составит около 7 мВт, а при 20°С – 14 мВт. По этим двум точкам строим зависимость мощности термогенеза пчелы от температуры в покое.
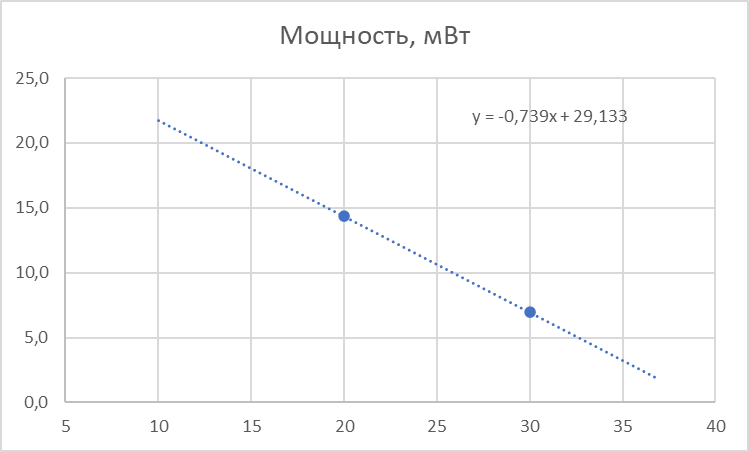
Таким образом, у пчелы есть “автономный подогреватель” никак не связанный с движителем полёта. Он работает постоянно, т.к. является продуктом основного обмена веществ, уровень которого меняется в зависимости от температуры окружающей среды. Вот эти два графика – и есть математическая модель пчелы в покое. Всё! Теперь мы можем для любой температуры воздуха (из рабочего диапазона) определить температуру пчелы и генерируемую ей тепловую мощность.
В полёте
В статье [3] приведены экспериментальные данные с измерениями температуры головы, торакса и брюшка в полёте в зависимости от температуры воздуха:
Регрессии, соответственно, следующие:

охлаждение испарением
Стоит обратить внимание на тот факт, что при температуре 28°С температура брюшка, а при температуре 35°С и температура головы ниже температуры воздуха. Этого пчела достигает за счёт испарения влаги при обдуве набегающим в полёте воздухом. Влагу она подаёт каплей в хоботок, а также открывая дыхальца, раздвигая тергиты. Человек и другие животные в жару тоже потеют чтобы не перегреться.
По этим экспериментальным данным построим тепловую математическую модель пчелы в полёте.
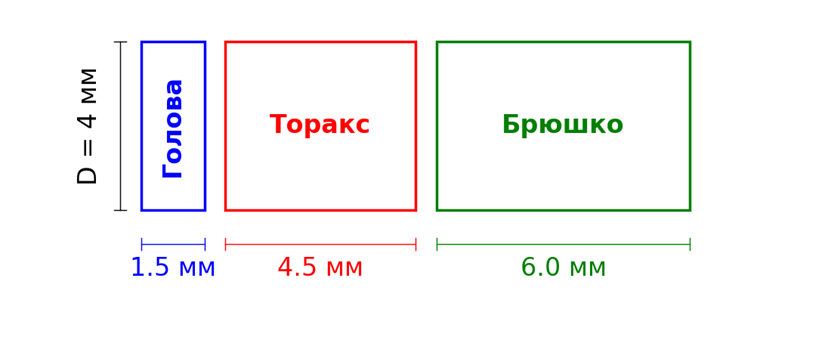
Представим пчелу в виде трех частей тела - головы (hd), торакса (th) и брюшка (ab), считая их цилиндрами диаметром 4 мм и высотой, соответственно, 1,5 мм, 4,5 мм и 6 мм. Они разделены/соединены небольшими зазорами и соединительными каналами. Площадь конвективного охлаждения (“обдува”) каждого цилиндра можно посчитать: головы - площадь переднего круга ![]() плюс площадь боковой поверхности головы
плюс площадь боковой поверхности головы ![]() , торакса - площадь боковой поверхности торакса
, торакса - площадь боковой поверхности торакса ![]() , брюшка - площадь боковой поверхности брюшка
, брюшка - площадь боковой поверхности брюшка ![]() плюс площадь заднего круга
плюс площадь заднего круга ![]() . Источник тепла находится в тораксе. Одна часть тепла рассеивается с поверхности торакса в воздух, другая - передаётся голове, третья - брюшку. В свою очередь голова и брюшко также рассеивают тепло со своей поверхности в воздух. Кроме того, голова и брюшко имеют возможность охлаждаться испарением воды через дыхальца и хоботок. Помимо контактного теплообмена (теплопроводность и конвекция) необходимо учесть теплообмен излучением. Система уравнений теплового баланса, таким образом, будет иметь вид:
. Источник тепла находится в тораксе. Одна часть тепла рассеивается с поверхности торакса в воздух, другая - передаётся голове, третья - брюшку. В свою очередь голова и брюшко также рассеивают тепло со своей поверхности в воздух. Кроме того, голова и брюшко имеют возможность охлаждаться испарением воды через дыхальца и хоботок. Помимо контактного теплообмена (теплопроводность и конвекция) необходимо учесть теплообмен излучением. Система уравнений теплового баланса, таким образом, будет иметь вид:

Тепловые сопротивления переходов торакс-голова “th-hd” и торакс-брюшко “th-ab” прямо пропорциональны средней величине зазора ![]() и обратно пропорциональны площади контакта с некоторыми коэффициентами пропорциональности
и обратно пропорциональны площади контакта с некоторыми коэффициентами пропорциональности ![]() и
и ![]() . Мощность, отводимая испарением, будем считать, определяется температурой торакса, т.к. именно его температуру пчеле нужно регулировать, именно там основной источник нагрева, т.е. она прямо пропорциональна температуре торакса с коэффициентами пропорциональности
. Мощность, отводимая испарением, будем считать, определяется температурой торакса, т.к. именно его температуру пчеле нужно регулировать, именно там основной источник нагрева, т.е. она прямо пропорциональна температуре торакса с коэффициентами пропорциональности ![]() и
и ![]() . Любое тело, имеющее температуру выше абсолютного нуля, излучает энергию - светится. Энергетическая светимость (Вт/м²) определяется формулой:
. Любое тело, имеющее температуру выше абсолютного нуля, излучает энергию - светится. Энергетическая светимость (Вт/м²) определяется формулой:

При температуре 12°С (285К) для абсолютно чёрного тела (![]() = 1) она составляет 375 Вт/м², а при 39°С - 538 Вт/м². Если умножить на площадь поверхности пчелы (2 см²), то получится, соответственно, 75 и 108 мВт. Однако пчела не абсолютно чёрное тело и не может излучать больше мощности метаболизма, коэффициент
= 1) она составляет 375 Вт/м², а при 39°С - 538 Вт/м². Если умножить на площадь поверхности пчелы (2 см²), то получится, соответственно, 75 и 108 мВт. Однако пчела не абсолютно чёрное тело и не может излучать больше мощности метаболизма, коэффициент ![]() определяет реальный уровень излучения. Кроме того, пчёлы находятся в окружении других предметов, они также имеют температуру выше абсолютного нуля и излучают. Пчела поглощает это излучение с таким же коэффициентом
определяет реальный уровень излучения. Кроме того, пчёлы находятся в окружении других предметов, они также имеют температуру выше абсолютного нуля и излучают. Пчела поглощает это излучение с таким же коэффициентом ![]() . Поэтому тепловое излучение обычно считают, как разность отдаваемой и поглощаемой мощности:
. Поэтому тепловое излучение обычно считают, как разность отдаваемой и поглощаемой мощности:

Как видно из этой формулы, при равенстве температур тела и среды теплообмена нет. Если температура тела выше температуры среды, то оно отдаёт тепло, если ниже - поглощает. Мощности теплового обмена излучением для принятой модели пчелы можно определить как:

Будем считать, что солнечный свет падает перпендикулярно продольной оси пчелы, тогда для принятых в модели размеров ![]() ,
, ![]() ,
, ![]() . Коэффициент поглощения полагаем одинаковым для торакса, головы и брюшка.
. Коэффициент поглощения полагаем одинаковым для торакса, головы и брюшка.
Значение коэффициента ![]() для пчелы неизвестно. Ранее я упомянул, что температура пчелы на солнце на 4°С выше, чем в тени. Увеличение температуры на
для пчелы неизвестно. Ранее я упомянул, что температура пчелы на солнце на 4°С выше, чем в тени. Увеличение температуры на ![]() вызовет увеличение конвекционных потерь, которые уравновесят тепло, полученное от прямых солнечных лучей:
вызовет увеличение конвекционных потерь, которые уравновесят тепло, полученное от прямых солнечных лучей:
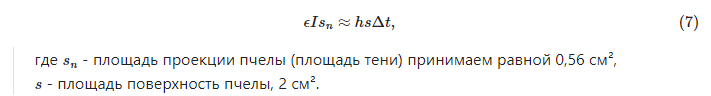
Из (7) находим ![]() . Это существенно ниже, чем обычно принято считать.
. Это существенно ниже, чем обычно принято считать.
Таким образом, задача создания модели, повторяющей данные экспериментов (уравнения (2)) сводится к подбору коэффициентов в системе уравнений(3). Их значения зависят от температуры воздуха. Для того, чтобы уменьшить размерность задачи начинаем с самой низкой температуры, при которой пчёлы начинают летать: 12°С. Полагаем, что при столь низкой температуре пчеле нет необходимости пользоваться механизмом испарения - она вряд ли будет потеть, поэтому можно коэффициенты ![]() и
и ![]() положить равными нулю. Также для начала положим, что прямого солнечного излучения на пчелу не падает - пасмурная погода, и I =0. Из этих условий подбираем в (3) коэффициенты
положить равными нулю. Также для начала положим, что прямого солнечного излучения на пчелу не падает - пасмурная погода, и I =0. Из этих условий подбираем в (3) коэффициенты ![]() и
и ![]() должны быть равны 0,065 и 0,042 - сопротивление к голове получилось больше, значит тепла туда передается меньше, чем брюшку. При этом средний зазор должен быть 0,68 мм. Точно так же (но уже учитывая возможность испарения и зафиксировав
должны быть равны 0,065 и 0,042 - сопротивление к голове получилось больше, значит тепла туда передается меньше, чем брюшку. При этом средний зазор должен быть 0,68 мм. Точно так же (но уже учитывая возможность испарения и зафиксировав ![]() и
и ![]() ) подбираем коэффициенты для других значений температуры воздуха: 15, 20, 25, 30 и 35°С. В итоге по этим точкам находим зависимости от температуры воздуха:
) подбираем коэффициенты для других значений температуры воздуха: 15, 20, 25, 30 и 35°С. В итоге по этим точкам находим зависимости от температуры воздуха:
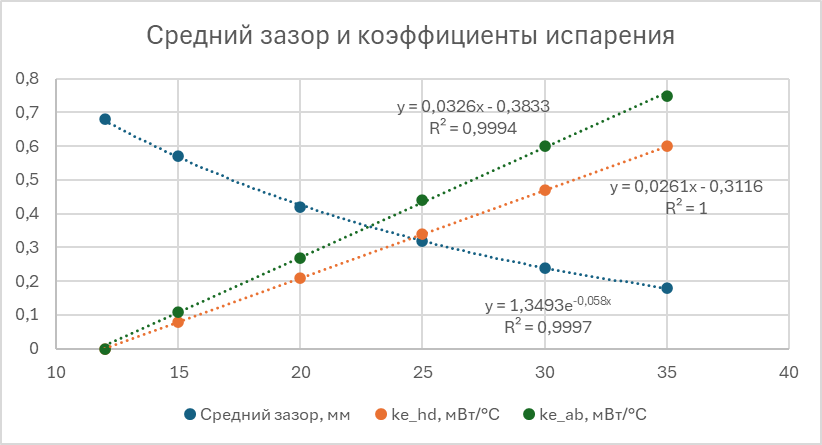
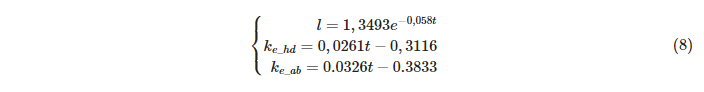
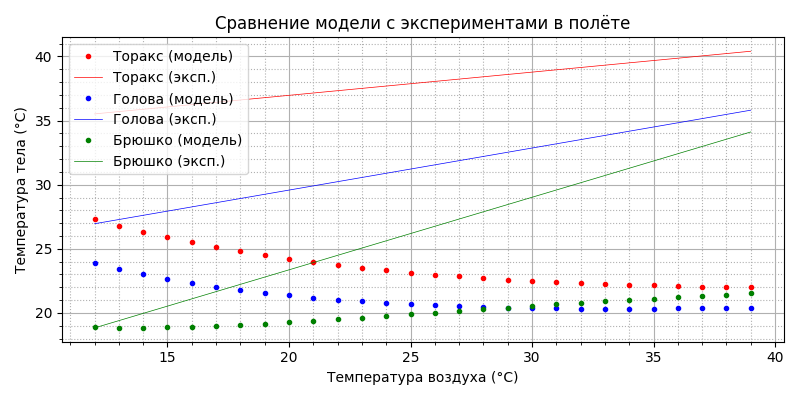
Подставляя полученные значения в модель теплового баланса, находим зависимости температуры торакса, головы и брюшка от температуры воздуха в модели и сравниваем с экспериментальными данными:
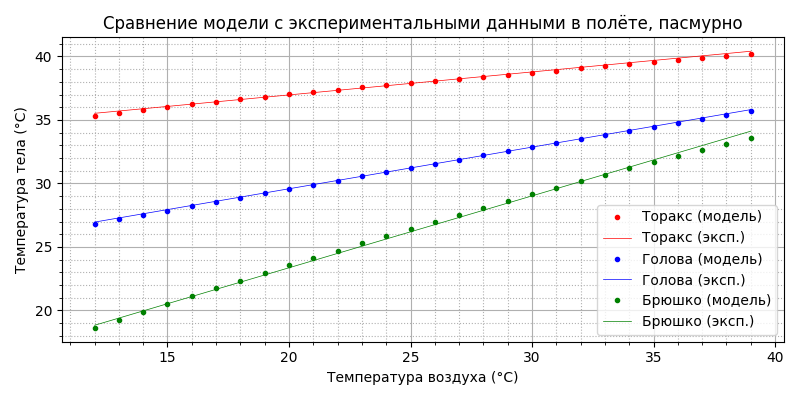
Как видим, совпадение неплохое. Напомню, эксперименты проводились в лабораторных условиях для парящего (машущего) полёта. Однако, для горизонтального свободного полёта требуемая мощность движителя примерно в два раза меньше, соответственно меньше будет и тепловая мощность. Посмотрим, что получится, если мощность термогенеза просто уменьшить вдвое:
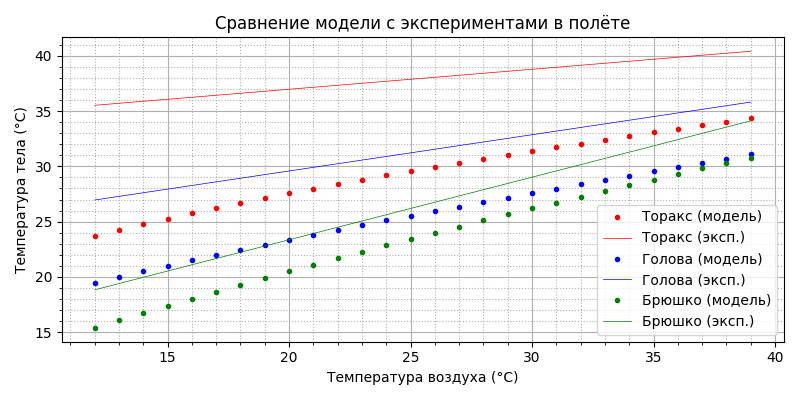
Пчела так не полетит - при температуре воздуха до 19°С торакс не прогревается до минимально необходимых 27°С, т.е. термогенеза недостаточно. Вспоминаем, что у пчелы же есть “автономный подогреватель”, который мы обнаружили при рассмотрении термогенеза “в покое”! Давайте посмотрим, что будет, если добавить термогенеза мощностью 29,133 - 0,739×T мВт (формула на графике мощности от температуры):
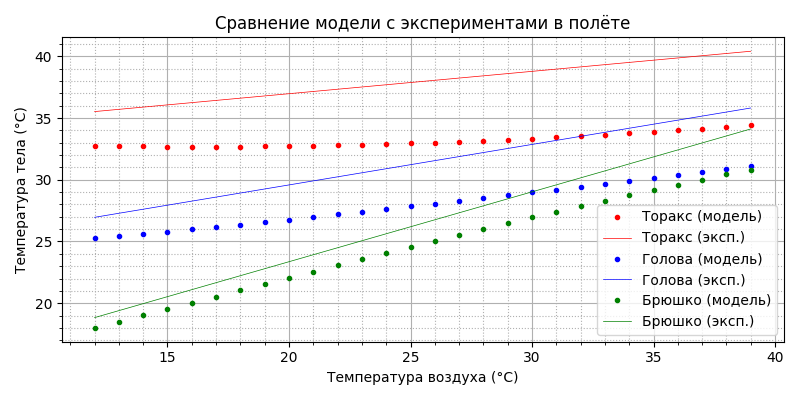
Да, видимо из-за охлаждения в горизонтальном полете пчела действительно догревает себя, как и в покое, тогда как при парящем полете мощность метаболизма и так близка к максимальной. Полученный результат заслуживает внимания: пчела способна стабилизировать температуру торакса в полёте во всём рабочем диапазоне: при изменении температуры воздуха от 12 до 39°С температура торакса (красные точки) меняется чуть больше, чем на 1 градус. В полёте пчела - теплокровное животное! При этом, начиная примерно с 34°С, температура торакса ниже температуры воздуха, брюшка - с 24°С, головы - с 29°С.
эксперимент с реальными пчелами-фуражирами
Справедливости ради стоит отметить, что ![]() - это приблизительная оценка для частного случая, и для разных пород и климатических условий коэффициент излучения может меняться в широких пределах, например, для темной пчелы он может быть, например, 0.9. Есть данные экспериментов 1985 года, проведенных в пустыне Сонора на реальных пчелах-фуражирах [4]:
- это приблизительная оценка для частного случая, и для разных пород и климатических условий коэффициент излучения может меняться в широких пределах, например, для темной пчелы он может быть, например, 0.9. Есть данные экспериментов 1985 года, проведенных в пустыне Сонора на реальных пчелах-фуражирах [4]:
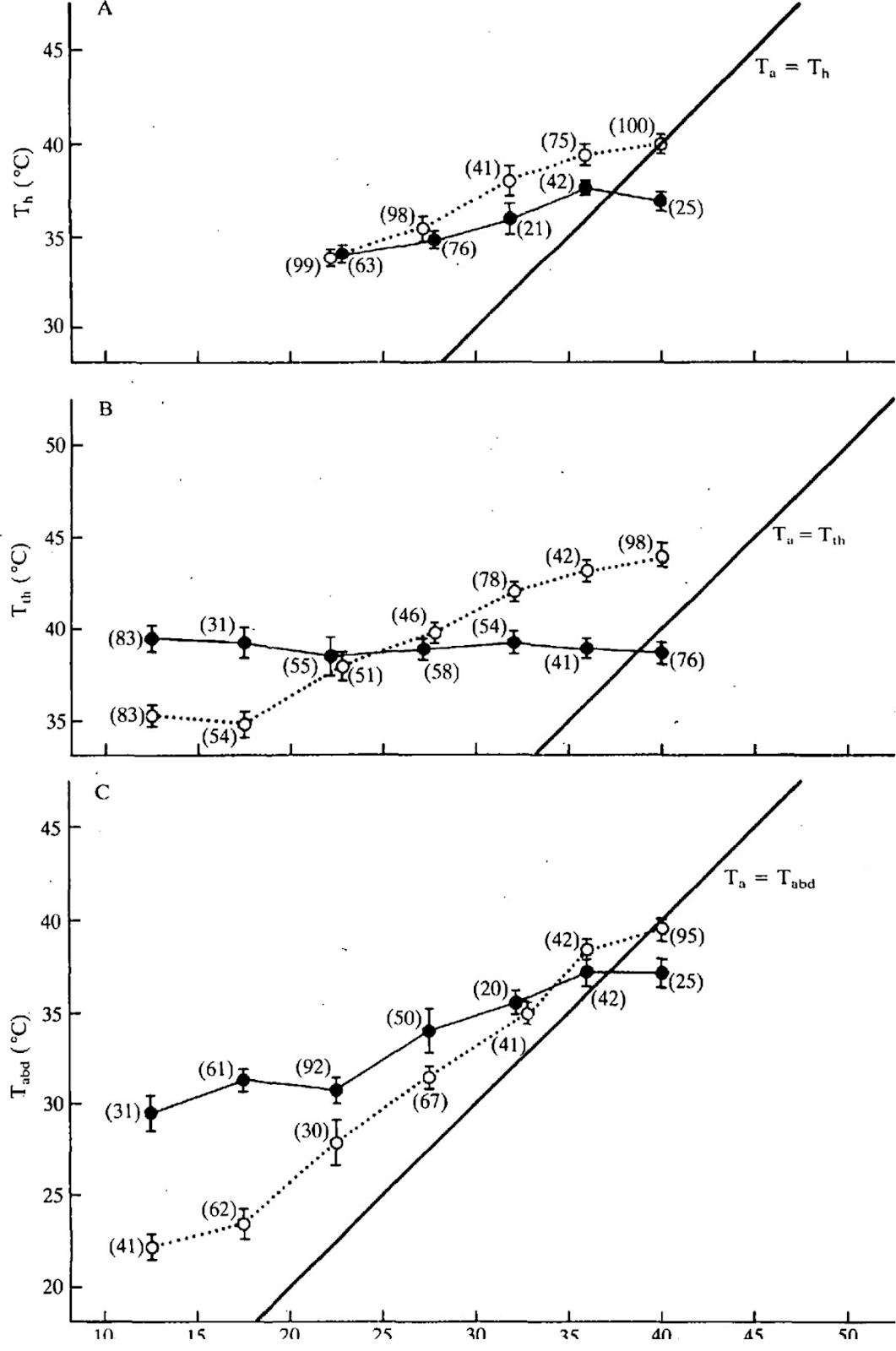
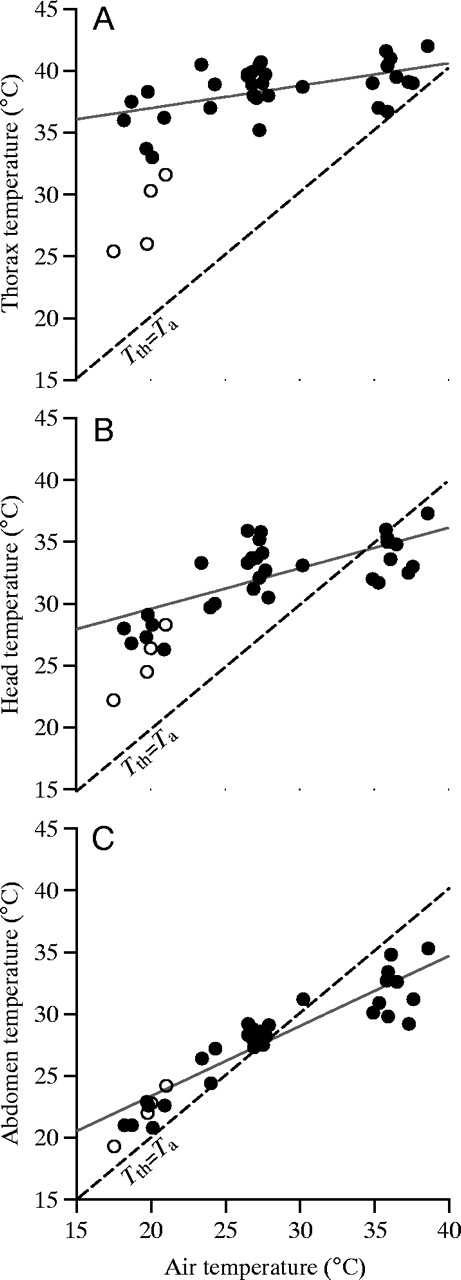
На рисунке показаны зависимости температуры торакса, головы и брюшка от температуры воздуха; сплошные линии - при полете “туда”, точечные - “обратно”. Видно, что температура торакса при полете “туда” поддерживается около 39-40°С, что возможно, например, при максимальной освещенности 1400 Вт/м² и ![]() или при освещенности 1200 Вт/м² и
или при освещенности 1200 Вт/м² и ![]() . Также на рисунке видно, что термостабилизация (теплокоровность) характерна лишь для полета “туда”, и пчела не заботится о ней при полете “оттуда”, пока температура торакса не опустится ниже 35°С.
. Также на рисунке видно, что термостабилизация (теплокоровность) характерна лишь для полета “туда”, и пчела не заботится о ней при полете “оттуда”, пока температура торакса не опустится ниже 35°С.
Таким образом, можно считать математическую модель термогенеза пчелы в полете построенной. Что на ней можно посмотреть?
Эксперименты на модели “в полете”
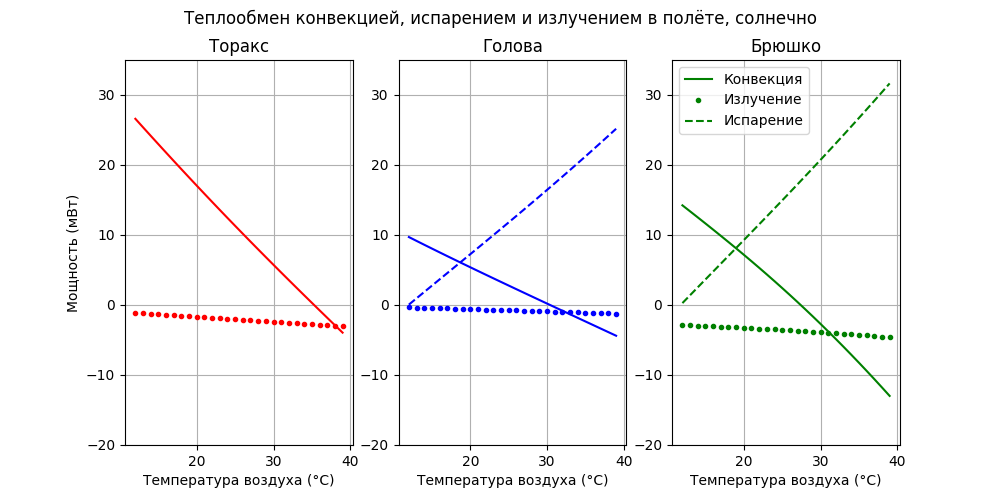
Посмотрим на модели как распределяется рассеиваемая мощность по частям тела пчелы:
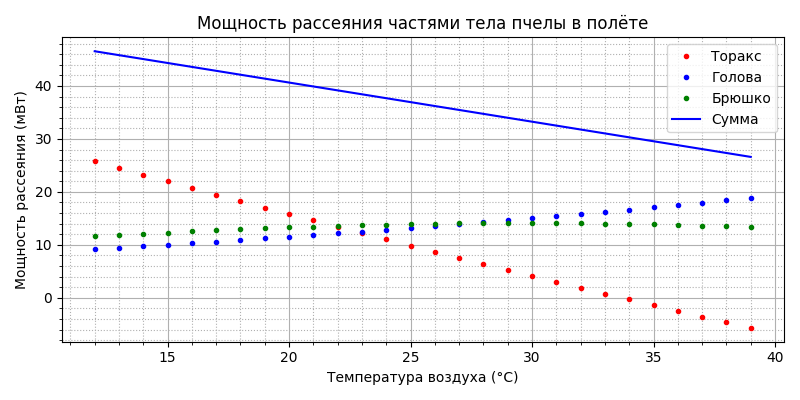
Из рисунка видно, что до 22°С основным “радиатором охлаждения” является торакс, но с ростом температуры воздуха величина рассеиваемой мощности снижается практически линейно от 26 мВт до нуля к 34°С, а затем становится отрицательной - т.е. нагревается воздухом. Тепло, отдаваемое брюшком остаётся практически постоянным в районе 12-14 мВт, головой - практически линейно растёт от 9 до 19 мВт. Общая рассеиваемая тепловая мощность снижается от 46 мВт при 12°С до 27 мВт при 39°С.
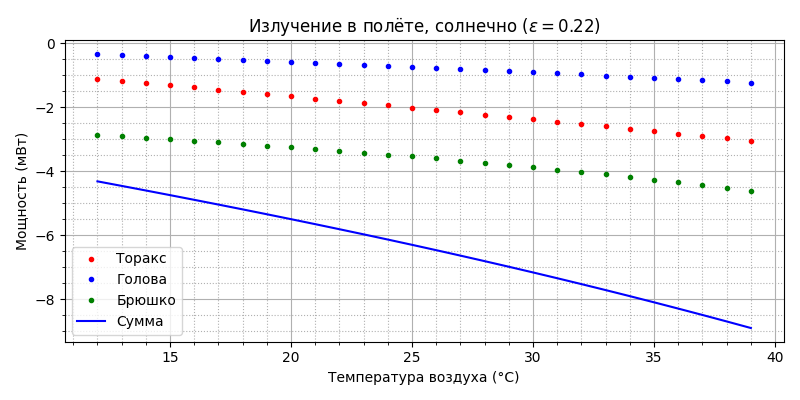
А вот как пчела излучает:
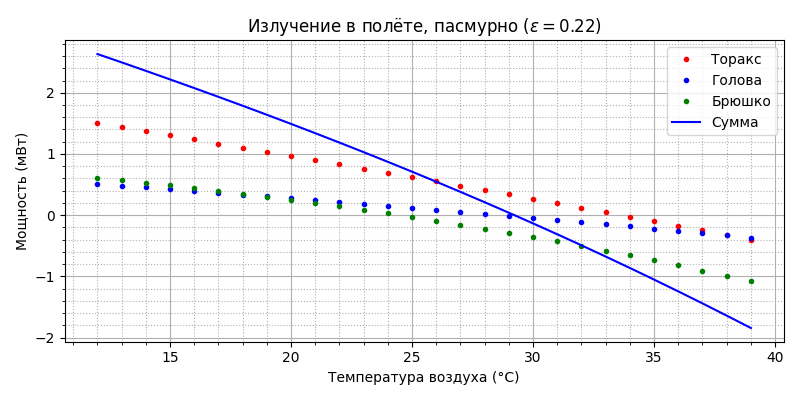
Видно, что вклад излучения пчелы в общий тепловой баланс незначительный, но до 29°С пчела излучением теряет, а после - поглощает тепло. Посмотрим как распределяется охлаждение на конвекцию, испарение и излучение:
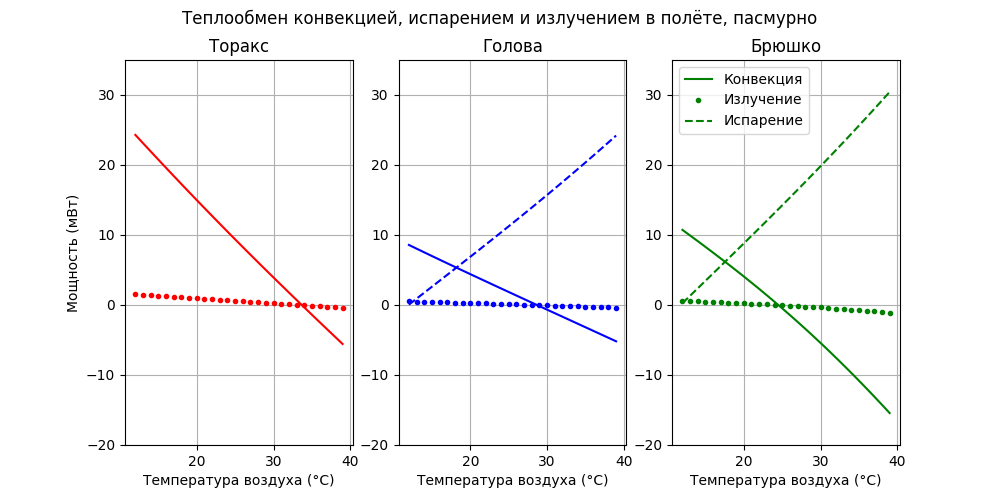
Как и ожидалось, с ростом температуры (после 18°С) основную роль в охлаждении играет испарение. Обдув после 24°С для брюшка и 29°С - для головы уже не охлаждает, а нагревает.
Рассмотрим теперь солнечный день (положим I = 700 Вт/м²).
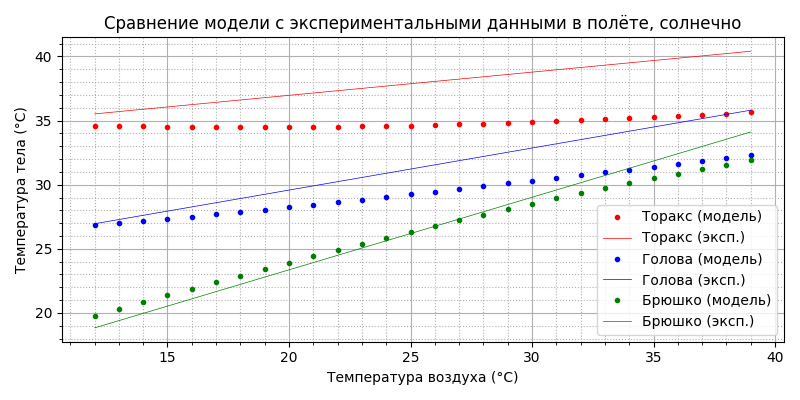
Видим, что температура торакса также стабильна, поднялась примерно на 2°С и составила 35-36°С, что, видимо, является наиболее типичным (пчёлы чаще всего работают в солнечную погоду) и комфортным для пчелы состоянием. Уровень теплообмена, несмотря на повышение температуры тела, остался примерно таким же, что и в пасмурную погоду:
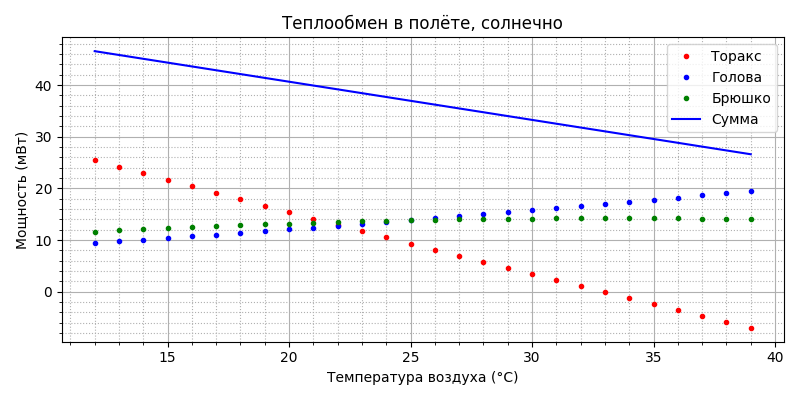
Это довольно неожиданно, но всё становится понятным, если посмотреть составляющие теплообмена:
охлаждение испарением осталось примерно таким же, а нагрев солнцем компенсировался конвекционным охлаждением (оно, напомню, пропорционально разности температур тела и среды).
Излучение отрицательное:
Это означает, что пчела в полёте подпитывается от солнца с мощностью от 4,5 до 9 мВт.
Можно, наверное, провести и другие модельные эксперименты.
Но давайте вернёмся на шаг назад. А работает ли предложенная модель для пчелы в состоянии покоя? Там мы представляли пчелу как единое тело. Что если и здесь рассмотреть усложнённую модель из торакса, головы и брюшка?
В покое. Итерация 1.
Положим в системе (3) ![]() , мВт, что соответствует состоянию “в покое”.
, мВт, что соответствует состоянию “в покое”.
Видно, что модель не подошла - на “одноэлементной” модели температура росла (по формуле (1)), а тут - падает. Но падает-то она от охлаждения испарением, а без полёта оно не эффективно - нет потока воздуха, “отрывающего” молекулы воды. Положим ![]() и
и ![]() равными нулю и вместо экспериментальных прямых нарисуем исходный график по формуле (1):
равными нулю и вместо экспериментальных прямых нарисуем исходный график по формуле (1):
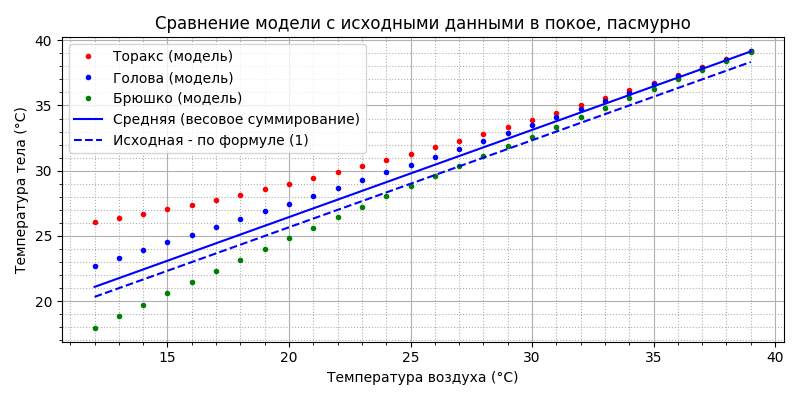
Действительно, при нулевом испарении наклон средней температуры в точности соответствует исходному по формуле (1) и испарению нужно снять всего 1°С, значит оно имеет постоянное значение и не зависит от температуры воздуха. Если задать ![]() и
и ![]() равными 0,03 мВт/°С, то получим
равными 0,03 мВт/°С, то получим
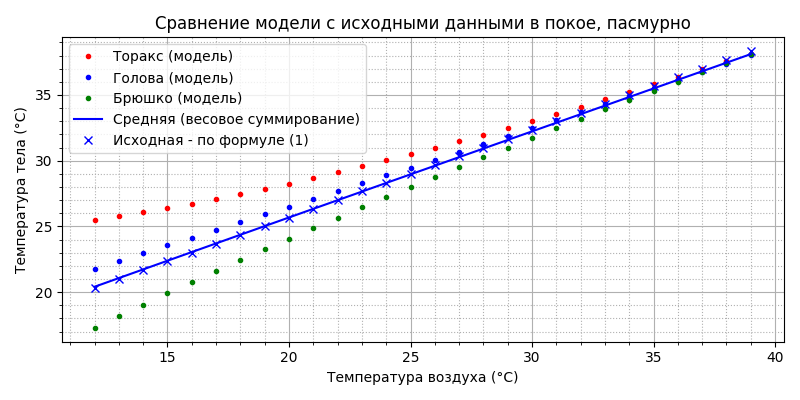
хорошее совпадение с исходными данными. Принципиально можно констатировать, что разработанная модель теплового баланса пчелы в полёте оказалась пригодна и для моделирования температуры пчелы “в покое”.
Эксперименты на модели “в покое”
Общий теплообмен:
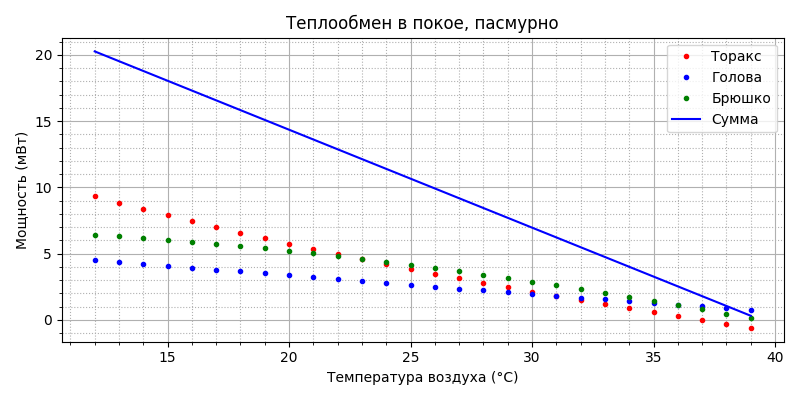
Видим, что в покое торакс также является основным радиатором, но выше 23°С больше отводит брюшко, а выше 31°С - и голова; после 36°С больше всего рассеивает голова. Замечаем также, что после 37°С торакс нагревается от воздуха. Общий уровень теплообмена примерно на 20 мВт меньше, чем в полете во всем рабочем диапазоне температур. Излучение:
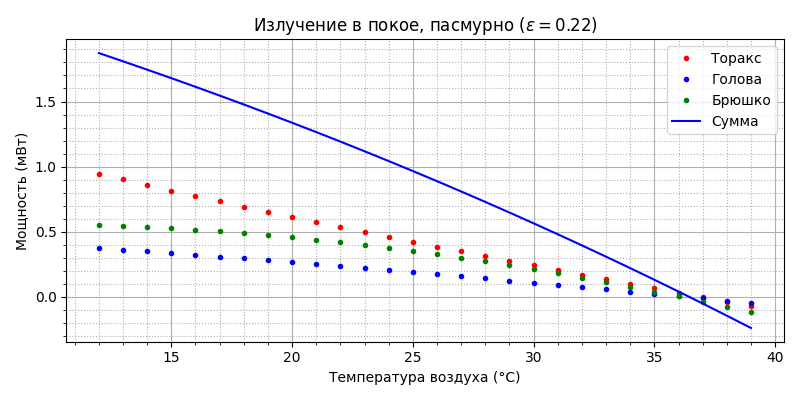
В покое в тени пчела отдаёт тепло излучением до температуры воздуха 36,5°С, а выше - получает; его уровень не превышает 10% от общего теплообмена.
Посмотрим, что в солнечную погоду. Температура:
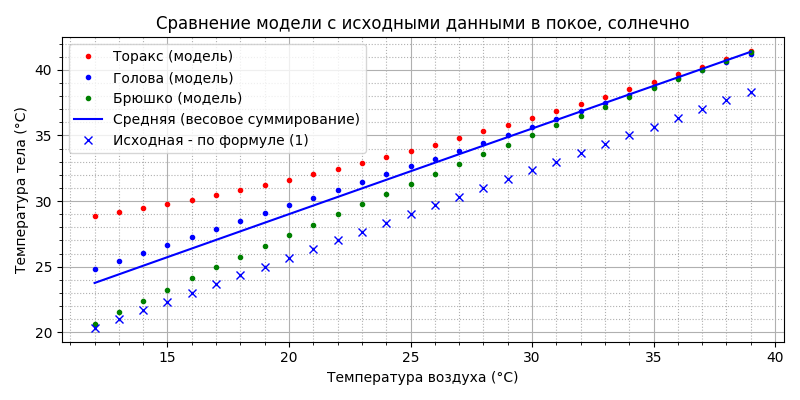
Видим, что, действительно, температура на 4°С выше, чем в тени. Теплообмен:
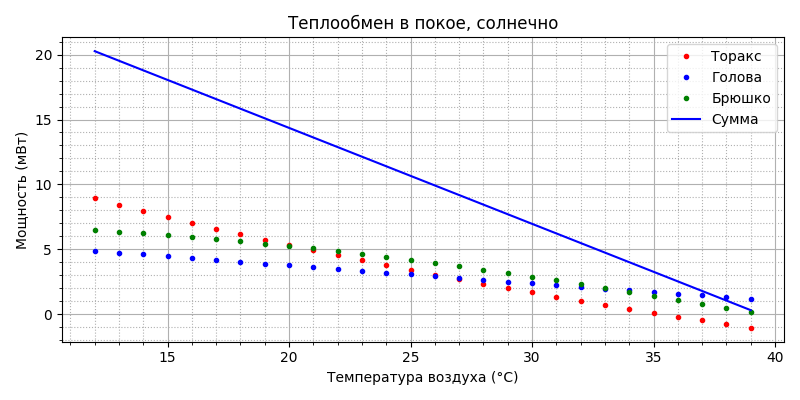
Как и в полёте, солнце практически не меняет скорость метаболизма. Излучение:
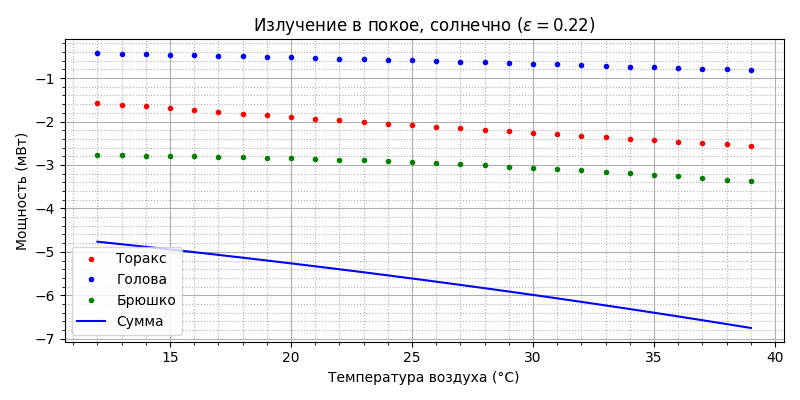
В покое, также как и в полёте, пчела с удовольствием подпитывается теплом от солнца с мощностью от 4,8 до 6,8 мВт. Результаты экспериментов выглядят вполне адекватными.
Режим печки
Пчеле для полёта необходима температура торакса не ниже 27°C. Однако, если мы посмотрим на график температуры пчелы в покое, то увидим, что это условие обеспечивается только при температуре воздуха выше 17°C в пасмурную погоду, а на солнце - выше 10°С. Но первые очистительные облёты пчёлы делают и в пасмурную погоду уже при температуре 10-12°C. Для того, чтобы взлететь пчела разогревает торакс до рабочей температуры путем изометрического сокращения летательных мышц. При этом махания крыльями не происходит. Такой режим можно назвать режимом печки. Это уже не стационарный (установившийся) режим - при разогреве температура изменяется во времени, поэтому уравнения теплового баланса будут выглядеть следующим образом:

де С_ - теплоёмкости соответствующих частей тела пчелы, а остальные обозначения те же, что и в системе (3). По ней же и вычисляются начальные условия - исходные температуры торакса, головы и брюшка при заданной температуре воздуха. Зададим температуру 12°C. Тогда исходные температуры будут, соответственно, торакса 25.47°C, головы 21.76°C, брюшка 17.28°C. Напомню, при этом ![]() , мВт - это зависящая от температуры воздуха мощность основного обмена - обычного метаболизма (автономного подогревателя). Если теперь добавить к этой мощности 45 мВт (мощность, близкая к максимальной для летательных мышц) , то переходной процесс будет выглядеть следующим образом:
, мВт - это зависящая от температуры воздуха мощность основного обмена - обычного метаболизма (автономного подогревателя). Если теперь добавить к этой мощности 45 мВт (мощность, близкая к максимальной для летательных мышц) , то переходной процесс будет выглядеть следующим образом:
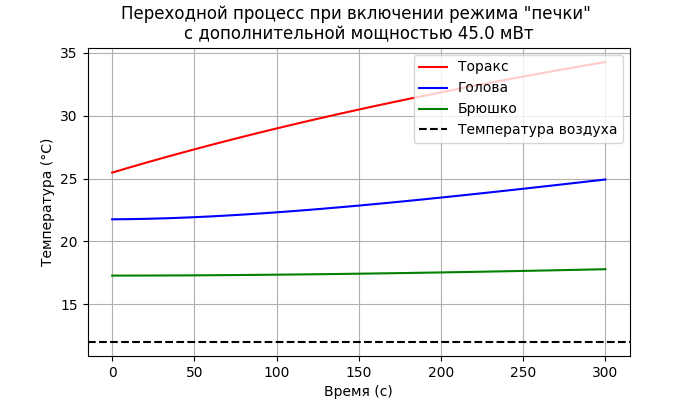
За две минуты температура торакса поднялась от 26 до 30°С, т.е. на 2°С в минуту, что соответствует известным экспериментальным данным.
Таким образом, в режиме печки пчела включает мощность близкую к максимальной - около 45 мВт. Этот же режим печки пчела включает, например, для обогрева расплода, или во время зимовки, когда надо поддержать температуру - мы к нему ещё вернёмся в следующей статье, которая будет о тепловом взаимодействии ячеек в пчелином соте. До ее публикации основные идеи можно посмотреть в моем блоге по пчеломоделированию.
Заключение: Разработана математическая модель физики теплообменных процессов в организме пчелы. Она основана на экспериментальных данных и не противоречит им.
Пчела имеет существенно разные температуры головы, торакса и брюшка; разница между температурой торакса и брюшка может доходить до 15°С, а торакса и головы - до 10°С при низких температурах; при высоких температурах разница меньше.
Для поддержания жизнедеятельности пчела расходует больше энергии метаболизма при низких температурах, и меньше - при высоких. Соответственно, при низких температурах она больше расходует корма.
Перед полётом в прохладную погоду пчела прогревает “двигатель”, включая режим печки мощностью около 45 мВт.
Термогенез и температура существенно отличаются в состояниях покоя и полёта - в полёте общий теплообмен больше - примерно на 20 мВт, что примерно соответствует мощности, расходуемой на горизонтальный полет (см. первую статью). При этом в полёте существенную роль в поддержании температуры играет испарение, коэффициент пропорциональности температуре торакса растёт с ростом температуры воздуха. В покое роль испарения значительно меньше, и коэффициент пропорциональности - постоянный. Также температура всех частей тела возрастает в солнечную погоду: в покое примерно на четыре градуса, в полёте - на два.
В полёте температура торакса слабо зависит от температуры окружающего воздуха, пчеле важно обеспечить стабильные условия работы своего движителя. В солнечную погоду - на уровне 35-36°С (что, видимо, является типичным и комфортным для пчелы режимом), в пасмурную - 32-33°С.
Разработанная модель позволяет оценить раздельно вклад различных механизмов - конвекции, испарения и излучения - в общий процесс теплообмена.
Литература
Еськов Е.К. Микроклимат пчелиного улья и его регулирование. М.: Россельхозиздат, 1978г., 79с.
Линдауэр М. Процессы регулирования в сообществах насекомых // Процессы регулирования в биологии: Перевод с немецкого.- М. Изд-во иностр. лит., I960.- С.178-195.
William A. Woods, Jr, Bernd Heinrich, Robert D. Stevenson: Honeybee flight metabolic rate: does it depend upon air temperature? - J Exp Biol (2005) 208 (6): 1161–1173.
Cooper, P. D., Schaffer, W. M. and Buchmann, S. L. (1985). Temperature regulation of honey bees (Apis mellifera) foraging in the Sonoran desert. J. Exp. Biol. 114, 1-15.