生物网络:
上面的这些内容整体上可以组成一个「图」,图中可以存在加权重的连接等等.
一维正反馈效应:
d[x]dt=v[x]nKdn+[x]n−d⋅[x]\frac{\text{d}[x]}{\text{d}t}=v\frac{[x]^n}{K_d^n+[x]^n} - d\cdot[x]
这个方程很明显难以得到解析的解,而且其解析解的实际意义也并不大. 事实上,我们更倾向于考虑分析这个方程的稳定性. 也就是对于 dx∗/dt=f(x∗)\text{d}x^*/\text{d}t=f(x^*) 这样的情况,分析 x=x∗+δxx = x^*+\delta x 造成的结果. 稳定态和不稳定态之间存在「分叉」.
提示
我认为,这里的一维方程分叉和理论力学混沌理论中的 Logistic 映射可能有类似的机制.
基因开关:考虑某个基因被抑制的概率为
pb(c)=cncn+Kdnp_b(c)=\frac{c^n}{c^n+K_d^n}
(仍然取一个 Hill 反应的形式.) 转录对应的方程为
dudt=−u+α1+vn,dvdt=−v+α1+un\frac{\text{d}u}{\text{d}t} = -u+\frac{\alpha}{1+v^n},\quad\frac{\text{d}v}{\text{d}t}=-v+\frac{\alpha}{1+u^n}
一共有两个基因,这是无量纲化的方程. 我们期望先找到不动点,也就是系统长时演化之后的情况;之后在不动点附近展开.
u0=α1+v0n,v0=α1+u0n⟹ddt(δuδv)=(fufvgugv)(δxδy)u_0=\frac{\alpha}{1+v_0^n},\quad v_0=\frac{\alpha}{1+u_0^n}\Longrightarrow\frac{\text{d}}{\text{d}t}\begin{pmatrix} \delta u\\\delta v \end{pmatrix}=\begin{pmatrix} f_u&f_v\\g_u&g_v \end{pmatrix}\begin{pmatrix} \delta x\\\delta y \end{pmatrix}
这个系数矩阵 (Jacobi Matrix) 给出两个本征值,它们的正负表现了两个不动点的稳定性,可能出现鞍点、旋进或者旋出的不同情况.
对于一个非常复杂的大型生物网络,我们的处理哲学是「Less is More」,期望达到的效果是把不重要的成分全部丢掉. 操作中类似于统计方法中的 kk - NN,用周围的状态给这个网格节点投票,决定其下一个步骤的演化.
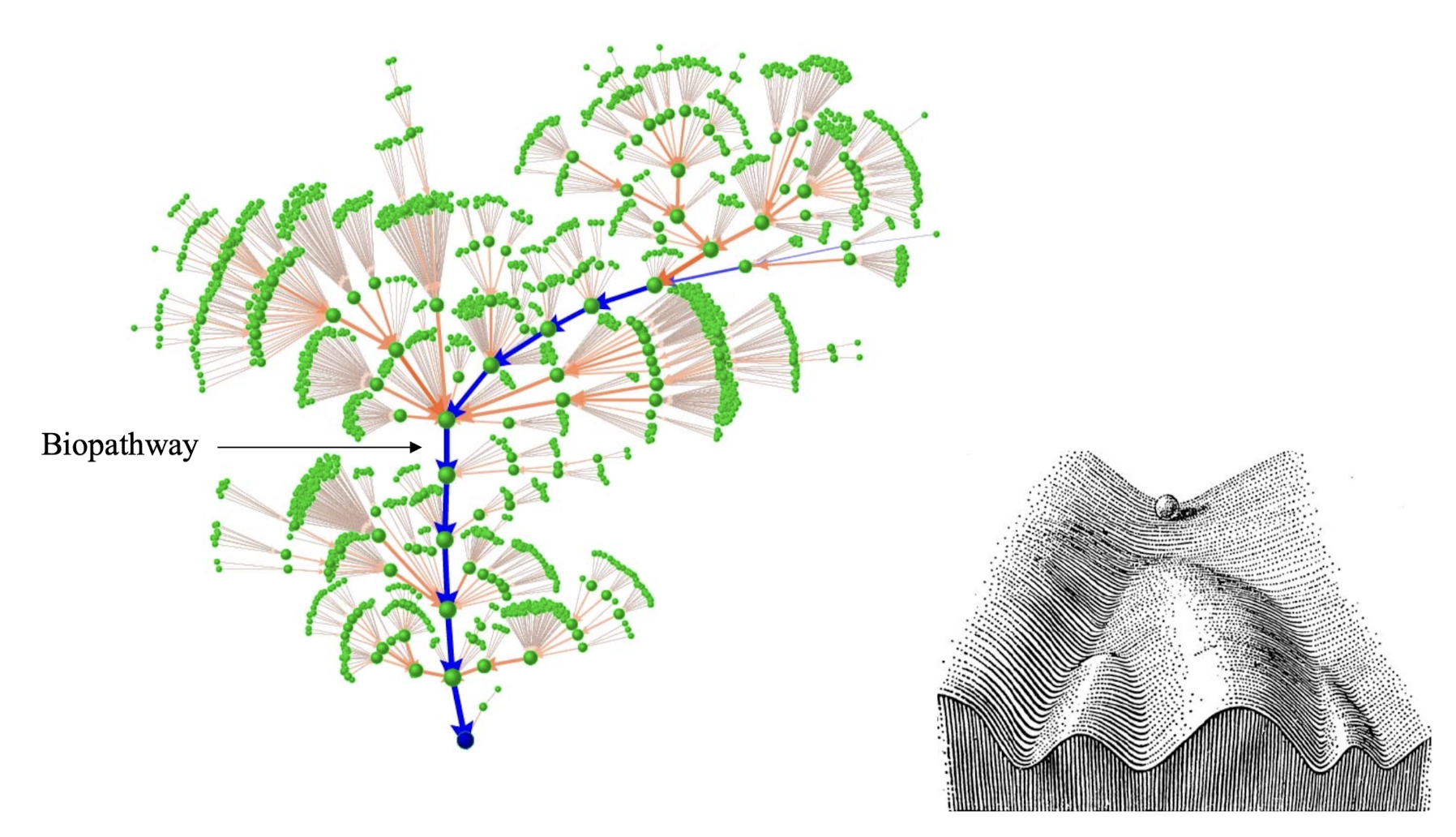
这样的主干道径迹就是一种所谓的吸引子,对于生物而言,甚至可以是一个吸引盆,生物网络的稳定性是非常好的. 这种稳健的特性本质上来源于网络自身的连接方式.
遗憾的是这样的理论取得的成果并不好... 因为我们并没有通过这个理论来解答很多实际问题,这是一个悲伤的故事;不过这样的事情也是常见的.
Hopfield Network & Spin-glass (自旋玻璃):
注意
物理学家在做什么,什么就是物理.
Hopfield 一开始仅仅是组建了一个二十多人的 seminar 团队而已,研究有没有什么 toy model 能够解释「记忆是如何储存在神经网络中的」. 这里面有神经科学家、动物行为学家,还有凝聚态物理学家... 非常混乱的一个团队. 但是也在这种情况下诞生了 Nobel 级别的理论.
自旋玻璃在某种意义上和 Ising 模型是类似的.
本学期课程的最后一个板块 —— 自组织.
0fcc8-feat(note): add note于 此内容由惯性聚合(RSS阅读器)自动聚合整理,仅供阅读参考。 原文来自 — 版权归原作者所有。